Centromere formation is a complicated process controlled by both genetic and epigenetic elements. Centromere function is conserved among different species and centromeric specific histone H3 variant CENH3 is also conserved, but the DNA sequences binding CENH3 nucleosomes change greatly. There are several megabases of repeat sequences in the centromere regions of plants. How does the centromere form?What factors decide CENH3 nucleosomes loading to centromere other than somewhere else? Does some specific DNA motif exist in centromere regions guiding CENH3 nucleosomes reposition? How are the retrotransposons and tandem repeat sequences arranged in the centromeric region? What’s the relation between DNA sequence and centromere function?
The group of Prof. HAN Fangpu from the Institute of Genetics and Developmental Biology, Chinese Academy of Science, studies centromere inactivation and
de novo centromere formation. The work of
de novo centromere sDic15 derived from nondisjunction and Breakage Fusion Bridge cycle of a B-A translocation chromosome revealed the role of DNA methylation and DNA motif in centromere formation, published in
The Plant Cell (DOI:
1105/tpc.113.110015), and the work of
de novo centromere on Dp3a produced by irradiation has been published in
Proc Natl Acad Sci USA (
DOI:10.1073/pnas.1303944110).
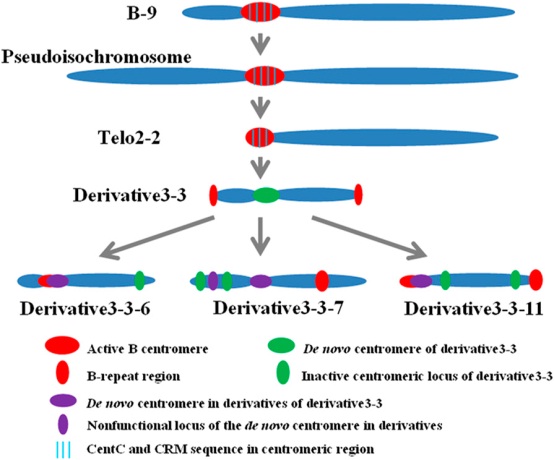
Recently, they found de novo centromere formation following centromere misdivision in maize. The misdivision of the B centromere on chromosome B-9 of TB-9Sb can produce derivatives with altered centromere sizes and centromeric DNA sequences. Among there variants, derivative 3-3 contains a de novo centromere following the loss of the functional B centromere of telo2-2 with retention of only reduced B-repeat copies. A 288 kb region from the short arm of chromosome 9 (9S) was involved in the centromere formation of derivative 3-3. Then, in the subsequent derivatives of 3-3, this de novo centromere was inactivated and other de novo centromeres in different regions were formed in the derivatives to take place of the centromere of 3-3.
This work suggested that the normal centromere must be large enough to be capable of purging any de novo centromeres that are initiated only in structurally acentric fragments, de novo centromeres become established because of the absence of such an inactivating force. In the sequence of events described here, it seems reasonable that the first formed de novo centromere in derivative 3-3 was insufficiently large to predominate over subsequently initiated de novo cases leading to inactivation and shifts to new functional sites of activity. Thus, despite regular initiation of small de novo centromeres, they are soon inactivated and chromosome structure is seldom altered over evolutionary time.
This work entitled “Sequential de novo centromere formation and inactivation on a chromosomal fragment in maize” has been published in Proc Natl Acad Sci USA on March 14, 2015 (DOI:10.1073/pnas.1418248112), with Ph.D candidates LIU Yalin and SU Handong as the co-first author. This research was supported by National Natural Science Foundation of China.
Figure. Centromere inactivation and de novo formation in maize chromosomal misdivision derivatives. The first derivative produced by misdivision of B-9 was known as a pseudoisochromosome (7). Then telo2-2 was derived from this pseudoisochromosome and derivative 3-3 derived from telo2-2 (23). Telo2-2 has a reduced B centromere. In derivative 3-3, the B centromere sequences are deleted but some B-repeat copies remain, and a 288-kb de novo centromere with no CentC and CRM sequence was formed. In three new derivatives of 3-3, the 288-kb centromere is inactivated. Another 200-kb de novo centromere has been formed in derivative 3-3-11.
Further rearrangements occurred in derivatives 3-3-6 and 3-3-11 with inactivation of the 288-kb site and a functional centromere apparently in the B chromosome sequences. (Image by IGDB)